
Equine Color Genetics
This page is a work in progress. Check back often.Click one of the genes below to jump to information on that gene or start from the beginning to learn about basic genetics first.
Colors:
Agouti (bay) ('A')
Champagne ('CH')
Cream ('CR')
Dun ('D')
Extension (black or red) ('E')
Pearl ('PRL')
Silver ('Z')
Agouti (bay) ('A')
Champagne ('CH')
Cream ('CR')
Dun ('D')
Extension (black or red) ('E')
Pearl ('PRL')
Silver ('Z')
Modifiers:
Flaxen ('F')
Gray ('G')
Pangare'('P')
Rabicano ('RA')
Roan ('RN')
Sooty ('STY')
Flaxen ('F')
Gray ('G')
Pangare'('P')
Rabicano ('RA')
Roan ('RN')
Sooty ('STY')
White Patterns:
Dominant White ('W')
Lethal White Overo (frame) ('O')
Sabino ('SB_')
Splash ('SPL')
Tobiano ('TO')
Leopard Complex and Patterns ('LP')
Dominant White ('W')
Lethal White Overo (frame) ('O')
Sabino ('SB_')
Splash ('SPL')
Tobiano ('TO')
Leopard Complex and Patterns ('LP')
Random:
Bend Or Spots
Birdcatcher Spots
Brindle
Lacing
Tetrarch Spots
Bend Or Spots
Birdcatcher Spots
Brindle
Lacing
Tetrarch Spots
To fully understand equine color genetics, one must first understand a few basic genetic principles. I will go over them first and hopefully give you a solid basis on which to build your genetic knowledge.
A gene is the smallest segment of DNA capable of producing a specific trait. A trait is defined as a distinct variant in phenotypic character of an organism. A simple example of a trait would be the white color of a flower. White in flowers is a genetically inherited characteristic, aka, a trait. A genotype is the genetic makeup of any given organism, whereas a phenotype is the outward appearance of the organism. We'll discuss how they relate to each other later on. For now we are going to take a close look at genotypes.
Each gene is made up of two alleles, one inherited from each parent. It is in this fashion that organisms reproduce in their likeness (a white flower produces another white flower). Each allele is passed at a 50% rate, ie allele1 (let's say white) is passed 50% of the time and allele2 (let's say red) is passed 50% of the time. When an organism has two identical alleles (two reds or two whites) it is said to be homozygous for that gene. If it carries one of each allele (one red and one white) it is considered heterozygous. More on what that means later.
So let's look at this from a more mathematical perspective. This is where the famous "punnet square" comes in. A punnet square is a diagram used to predict the outcome of a particular cross. Dominant alleles are generally denoted by a capital letter representing the gene the allele is "on" (the red gene for this example). In the case of red flowers vs. white flowers (using "simple dominance", which we are for now), red would be 'R' and white would be 'r'. This is a punnet square predicting the possible outcome of two flowers who are both heterozygous (thus carrying one each of the red and white alleles):
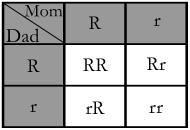
This square shows us that there are four possible genotypes for this particular gene mating. 25% will be homozygous for the red allele, 25% will be heterozygous by receiving a red allele from Mom and a white allele from Dad, 25% will be heterozygous by receiving a white allele from Mom and a red allele from Dad, and 25% will be homozygous for the white allele.
As you can see, often times each parent passes a different form (allele) of any given gene (ie one passes the white allele and the other passes the red allele) causing a bit of a conflict. Such disputes are solved by a set order of dominance. There are three different types of dominance. In equine color genetics you will need to know a bit about each type of dominance so I will give you some examples to work with.
Simple dominance: In flowers (for this example), red is dominant to white so the flower inheriting both a red and a white allele will be red. That is a classic example of simple dominance. That flower will be able to produce either red or white flowers depending on the genetics of his mate and is thus considered to be heterozygous for that gene. If the flower were grown with two red alleles instead it would still be red but it would be considered homozygous for the red allele. Due to the red allele's dominance, a flower carrying even one red allele will always be red. Any flower produced from a homozygous red parent will always be red regardless of the other parent's contribution as the homozygous parent only has red to pass. A flower homozygous for white can only pass the white allele so the color of any flowers created from it will ultimately be determined by the flower it is crossed with. Two flowers homozygous for the white allele can only create other white flowers. The same goes for homozygous reds of course. Let's look at a couple more punnet squares so you can see how different zygosity of the parent's can affect the resulting flower.
Now you can see how one parent being homozygous for a given allele will affect matters:
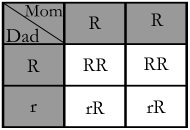
With Mom being homozygous for the dominant allele (red), you can see all flowers will inherit a dominant allele from her. In this case all of her flowers will be red with 50% of them inheriting Dad's recessive allele and having the ability to produce white flowers themselves.
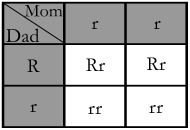
In this case Mom's being homozygous for the recessive allele means that each of her flowers will have the ability to produce white flowers but due to Dad's dominant allele, only 50% of them will be white themselves.
Incomplete dominance: Another form of dominance you will need to understand is the "incomplete dominant". In the case of an incomplete dominant the dominant allele has a different expression in the homozygous state. Whereas our flower which is heterozygous for red/white is red and our flower which is homozygous for red is also red, an incomplete dominant gene's expression would mean the flower would have a distinctly different appearance when comparing the heterozygote to the homozygote. In this example the heterozygous flower would be *pink*, an even blend of the two alleles, and the homozygous red flower would be red. The homozygous white flower would still be white. The inheritance of an incomplete dominant gene is the same, only the phenotypical expression varies.
When writing a punnet square for an incomplete dominant trait, it is better to use capital letters for both alleles, separating them by phenotype instead. We'll use R and W.

So here you can see that 25% will be red, 50% will be pink and 25% will be white.
Co dominance: In addition to simple dominance and incomplete dominance there is one more type of dominance that will impact your understanding of equine color genetics. Co-dominance in simple terms means that both alleles are dominant and they work together. Using our red and white flowers example, an RR flower would be red, WW would be white and RW would be *striped*, a combination of the two alleles but not a blend like the incomplete dominant. Red and white share equal turf in this instance instead of melding together. The inheritance of a co-dominant gene is also the same, only the expression differs.

So here you can see that 25% will be red, 50% will be striped and 25% will be white.
Now that you probably have a good understanding of the basics of genetic inheritance, we'll move on to the specific equine color genes and their impact on the equine phenotype.
Click on the photos to see the full size image. When you are finished click the "BACK" button to return to this page.
")
") Extension 'E' and 'e' (simple dominant): Extension is the basis of all equine color and the most important gene of all. The dominant allele of extension 'E' causes the production of eumelanin in the horse's coat causing a "black base color". The recessive allele of extension causes the production of pheomelanin in the horse's coat causing a "red base color". Every horse has either a black base or a red base. ALL other phenotypes other than red and black are caused by the addition of other modifiers/diluters. Read on to learn about those.
Extension 'E' and 'e' (simple dominant): Extension is the basis of all equine color and the most important gene of all. The dominant allele of extension 'E' causes the production of eumelanin in the horse's coat causing a "black base color". The recessive allele of extension causes the production of pheomelanin in the horse's coat causing a "red base color". Every horse has either a black base or a red base. ALL other phenotypes other than red and black are caused by the addition of other modifiers/diluters. Read on to learn about those.EE = homozygous black. Ee = heterozygous black. ee = homozygous red.
")
") Agouti 'A' and 'a' (simple dominant): Agouti is typically referred to as the "bay gene". It restricts the production of eumelanin on the head and body of the horse leaving the mane, tail and legs in their original state. Because the gene only restricts eumelanin, it has no effect on red based horses. Red based horses can carry agouti and thus affect their offspring by passing it on, but they will not phenotypically differ from a red based horse without agouti. Agouti is believed to be a "multiple allelic series" meaning there are more alleles "on" the gene in addition to 'A' and 'a'. Each of the alleles produces a different phenotype and there is a theorized order of dominance among them. The other alleles are believed to be wild bay 'A+' (dominant over all the others), bay 'A' (dominant over the following two), brown 'At' (dominant over the next) and "recessive" 'a' (no impact on the horse's color). Each horse will still only have two alleles (one from each parent), but depending on which two are present the phenotype will vary.
Agouti 'A' and 'a' (simple dominant): Agouti is typically referred to as the "bay gene". It restricts the production of eumelanin on the head and body of the horse leaving the mane, tail and legs in their original state. Because the gene only restricts eumelanin, it has no effect on red based horses. Red based horses can carry agouti and thus affect their offspring by passing it on, but they will not phenotypically differ from a red based horse without agouti. Agouti is believed to be a "multiple allelic series" meaning there are more alleles "on" the gene in addition to 'A' and 'a'. Each of the alleles produces a different phenotype and there is a theorized order of dominance among them. The other alleles are believed to be wild bay 'A+' (dominant over all the others), bay 'A' (dominant over the following two), brown 'At' (dominant over the next) and "recessive" 'a' (no impact on the horse's color). Each horse will still only have two alleles (one from each parent), but depending on which two are present the phenotype will vary.")
 Champagne 'CH' and 'ch' (simple dominant): Champagne is a dilution gene that affects both eumelanin and pheomelanin. Champagne foals are always born with blue eyes and pink skin. Generally champagne foals start out several shades darker than their adult color will be. Over time the blue eyes change to an amber color and the skin becomes mottled with small, dark freckles. Some champagne's mottling can become so extensive as to give the skin an overall gray/pink appearance, however a peek under the tail or near the udder or sheath should reveal very obvious pink skin with few freckles. Amber eyes and pink skin are must for a horse to be considered champagne. Some non-champagnes can also show these characteristics; though generally to a lesser extent (palominos are notorious for it). Champagne on a red based horse can vary from a very dark red/gold with a body colored mane and tail to a very light tan/gold with a white mane and tail and they are considered "gold champagne". They tend to look very similar to palominos but the mottled skin and amber eyes can generally clue you in to the genotype. Bay horses carrying the champagne typically looks very much like buckskins with slightly lighter and more brownish toned legs (of course the characteristics can help you here too) and are considered "amber champagne". Champagnes with a black base and considered "classic champagne" and are generally a reasonably dark and charcoal like color. Sometimes classic champagne can be confused with grullo (black dun) but the champagne will not show dun characteristics such as the dorsal stripe and leg barring, etc. When a horse carries a champagne and a cream gene together, "cream" is added to the name, ie "gold cream champagne".
Champagne 'CH' and 'ch' (simple dominant): Champagne is a dilution gene that affects both eumelanin and pheomelanin. Champagne foals are always born with blue eyes and pink skin. Generally champagne foals start out several shades darker than their adult color will be. Over time the blue eyes change to an amber color and the skin becomes mottled with small, dark freckles. Some champagne's mottling can become so extensive as to give the skin an overall gray/pink appearance, however a peek under the tail or near the udder or sheath should reveal very obvious pink skin with few freckles. Amber eyes and pink skin are must for a horse to be considered champagne. Some non-champagnes can also show these characteristics; though generally to a lesser extent (palominos are notorious for it). Champagne on a red based horse can vary from a very dark red/gold with a body colored mane and tail to a very light tan/gold with a white mane and tail and they are considered "gold champagne". They tend to look very similar to palominos but the mottled skin and amber eyes can generally clue you in to the genotype. Bay horses carrying the champagne typically looks very much like buckskins with slightly lighter and more brownish toned legs (of course the characteristics can help you here too) and are considered "amber champagne". Champagnes with a black base and considered "classic champagne" and are generally a reasonably dark and charcoal like color. Sometimes classic champagne can be confused with grullo (black dun) but the champagne will not show dun characteristics such as the dorsal stripe and leg barring, etc. When a horse carries a champagne and a cream gene together, "cream" is added to the name, ie "gold cream champagne".")
") Cream 'CR' and 'cr' (incomplete dominant): Cream is the wonderful dilution gene that gives us so many oh-so-desirable phenotypes. This gene only fully effects pheomelanin and thus has little to no effect on black pigment. Cream's effect on a horse's color is dependant on the horse's zygosity for the gene. While one copy of cream ('CRcr') is considered a single cream dilute and is a partial dilution of the horse's original color, two copies of cream ('CRCR') is known as a double dilute and dilutes the horse's coat almost completely (they generally look very close to white). One copy of cream on a red based horse is what we know as a palomino. They generally are some shade of gold with a white (or light) mane and tail. One copy of cream on a bay based horse gives us buckskin who are usually also some shade of gold with a black mane, tail and legs. The reason the points remain undiluted on a buckskin is due to cream's lack of effect on black pigment. The body and head of a buckskin is diluted because the agouti gene has already made those areas red. One copy of cream on a black based horse is called a smokey black and nearly always looks like a "normal black". Most of the time a smokey black can only be identified by a genetic test for the cream gene. Two copies of cream (and thus homozygous) on a red based horse is a cremello and generally look very close to white. Two copies of cream on a bay based horse is a perlino and it's body and head look nearly white while it's mane, tail and legs tend to have slightly more color and may appear to be a super light orange color. Cream in it's homozygous state does dilute black pigment to a significant extent thus the bay and black based homozygotes will usually show this light orange color where there black would have been. Two copies of cream on a black base is referred to as a smokey cream. Smokey creams have an overall body color very similar to the legs of the perlino, a very light orange, sometimes bordering on whitish.
Cream 'CR' and 'cr' (incomplete dominant): Cream is the wonderful dilution gene that gives us so many oh-so-desirable phenotypes. This gene only fully effects pheomelanin and thus has little to no effect on black pigment. Cream's effect on a horse's color is dependant on the horse's zygosity for the gene. While one copy of cream ('CRcr') is considered a single cream dilute and is a partial dilution of the horse's original color, two copies of cream ('CRCR') is known as a double dilute and dilutes the horse's coat almost completely (they generally look very close to white). One copy of cream on a red based horse is what we know as a palomino. They generally are some shade of gold with a white (or light) mane and tail. One copy of cream on a bay based horse gives us buckskin who are usually also some shade of gold with a black mane, tail and legs. The reason the points remain undiluted on a buckskin is due to cream's lack of effect on black pigment. The body and head of a buckskin is diluted because the agouti gene has already made those areas red. One copy of cream on a black based horse is called a smokey black and nearly always looks like a "normal black". Most of the time a smokey black can only be identified by a genetic test for the cream gene. Two copies of cream (and thus homozygous) on a red based horse is a cremello and generally look very close to white. Two copies of cream on a bay based horse is a perlino and it's body and head look nearly white while it's mane, tail and legs tend to have slightly more color and may appear to be a super light orange color. Cream in it's homozygous state does dilute black pigment to a significant extent thus the bay and black based homozygotes will usually show this light orange color where there black would have been. Two copies of cream on a black base is referred to as a smokey cream. Smokey creams have an overall body color very similar to the legs of the perlino, a very light orange, sometimes bordering on whitish.")
") Dun 'D' and 'd' (simple dominant): Dun is one of the most intriguing dilutions in my opinion and is my personal favorite. It creates a look on red and bay based horses very similar to cream and adds characteristics called "dun factor". Dun's *must* have a clear, defined dorsal stripe in order to be considered dun and having any of the others characteristics is sometimes considered required and always desirable: leg barring, wither shading, face masking, face cob webbing, ear tips, mane and tail guard hairs, leg marbling, neck shadows and dorsal barbs. These markings are the color the body of the horse would have been had it not been diluted by the dun gene's effects. Red based horses displaying the dun gene range from a light pinkish tone to a deep coppery red with darker legs, mane and tail and are called red dun. Dun diluted horses with a bay base look VERY much like buckskins most of the time with a diluted body and head and black legs, mane and tail and are called bay dun. Black based horses with the dun gene vary from a dark color very close to black to a very light grayish/charcoal appearance and normally show at least a dorsal stripe and leg bars and normally always have a darker head. They are called grulla or grullo (Spanish female and male pronouns describing a blue crane) but a more accurate name would be black dun.
Dun 'D' and 'd' (simple dominant): Dun is one of the most intriguing dilutions in my opinion and is my personal favorite. It creates a look on red and bay based horses very similar to cream and adds characteristics called "dun factor". Dun's *must* have a clear, defined dorsal stripe in order to be considered dun and having any of the others characteristics is sometimes considered required and always desirable: leg barring, wither shading, face masking, face cob webbing, ear tips, mane and tail guard hairs, leg marbling, neck shadows and dorsal barbs. These markings are the color the body of the horse would have been had it not been diluted by the dun gene's effects. Red based horses displaying the dun gene range from a light pinkish tone to a deep coppery red with darker legs, mane and tail and are called red dun. Dun diluted horses with a bay base look VERY much like buckskins most of the time with a diluted body and head and black legs, mane and tail and are called bay dun. Black based horses with the dun gene vary from a dark color very close to black to a very light grayish/charcoal appearance and normally show at least a dorsal stripe and leg bars and normally always have a darker head. They are called grulla or grullo (Spanish female and male pronouns describing a blue crane) but a more accurate name would be black dun.